Unidade VII - Bases genéticas do Cancêr
2. Genes Supressores Tumorais
Os supressores tumorais são genes que expressam produtos que controlam o ciclo celular (G1, S, G2, M) e quando mutados deixam de exercer seus papéis através de processos específicos para cada gene.
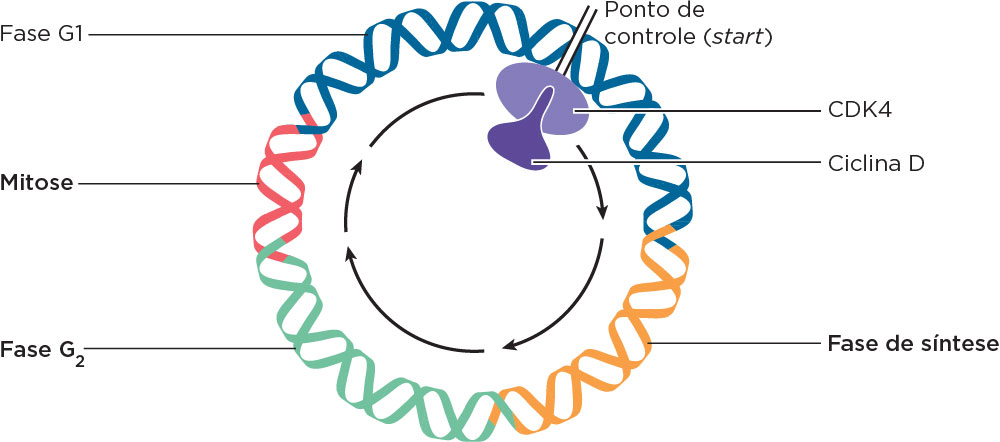
O ciclo celular apresenta períodos de síntese de proteínas específicas e DNA, e de divisão do material genético. Os ciclos são controlados por sinais externos ou extrínsecos à células, como fatores de crescimento e hormônios, e por sinais internos à celulas, principalmente por duas classes de proteínas: as ciclinas (nome proveniente da síntese e degradação ao final de um ciclo) e as proteínas quinases (com atividade de fosforilação de outras proteínas) dependentes de ciclinas chamadas de CDKs (ou Cdcs). As transições entre as diferentes fases do ciclo são controladas por “pontos de controle” (checkpoints) que o param, por exemplo, em resposta ao DNA danificado ou até que a síntese do DNA seja completada. A formação do complexo ciclinas-CDKs é um evento crítico nesse processo.
Um dos pontos de checagem mais importantes é chamado de start, que está entre a metade de G1 e S. Nesse ponto, a formação do complexo ciclinas-CDKs e a fosforilação de proteínas, como a pRB, é importante para a entrada na fase S. As células nas quais o start está alterado são propensas a se tornarem cancerosas, pois o DNA danificado deve ser reparado antes da replicação do DNA, com importante atuação da proteína pRB . O gene RB1 está situado no cromossomo 13 e foi descoberto por meio de mutações na pRB em retinoblastoma; por isso seu gene recebeu esse nome, mas se expressa em vários tecidos. A proteína produzida pelo RB bloqueia o ciclo celular quando não está fosforilada. Nesta forma, a proteína pRB se liga ao fator de transcrição E2F. Após a fosforilação, a pRB se separa de E2F que atuará na ativação de genes-alvo de progressão do ciclo celular. Quando o RB1 está mutado, seu produto encontra-se alterado podendo ser encontrado permanentemente fosforilado, não se ligando a E2F o que resultará na progressão do ciclo celular para a fase S e, por fim, a mitose. Assim, na ausência do freio por RB e de outros freios do ciclo celular, as células se dividem incessantemente formando os tumores. As mutações no gene RB têm sido descritas em vários cânceres como os de próstata, bexiga e osteossarcomas. Estudos em camundongos demonstraram que as mutações homozigotas herdadas nesse gene levam à morte logo após o nascimento .
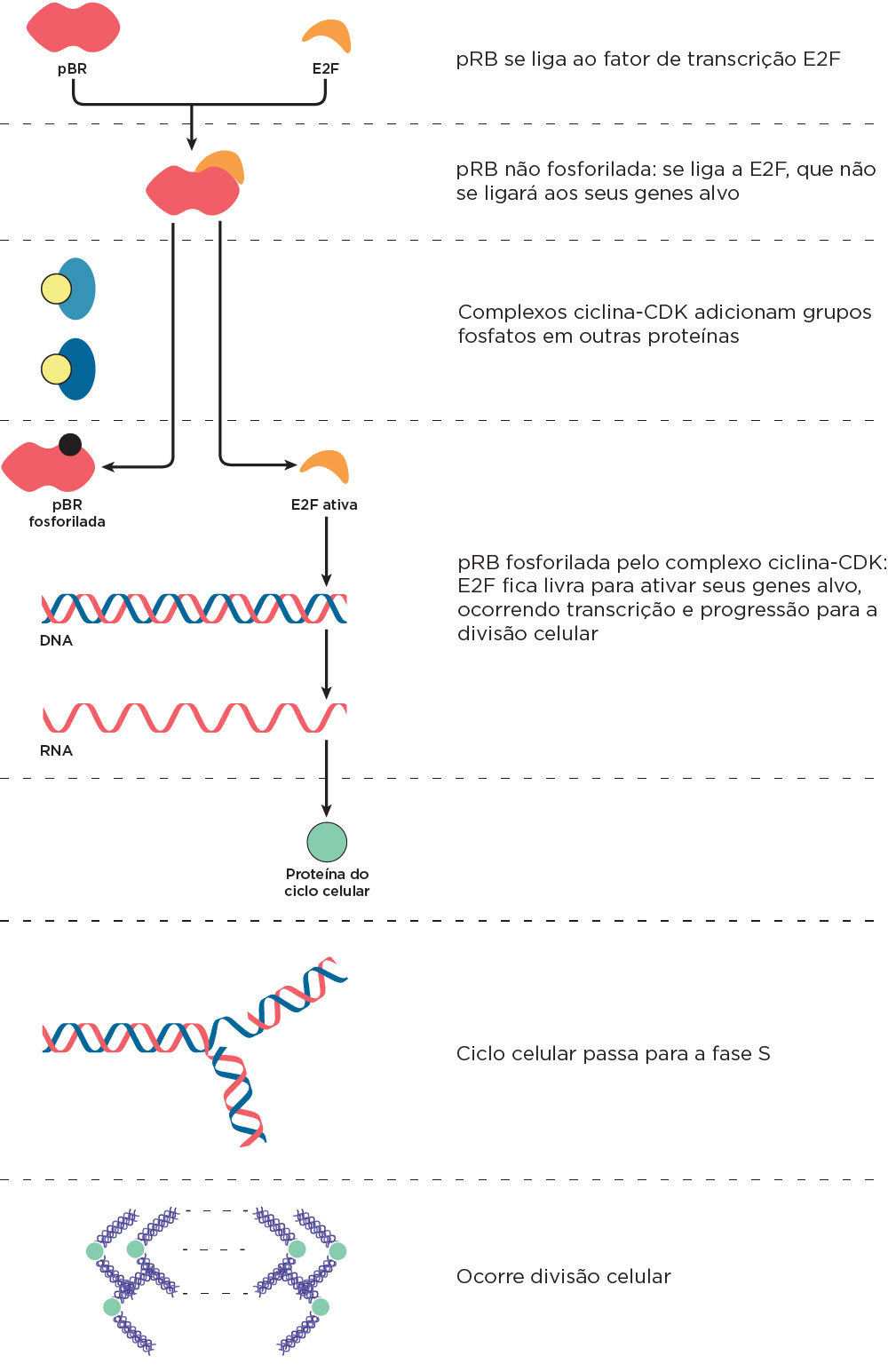
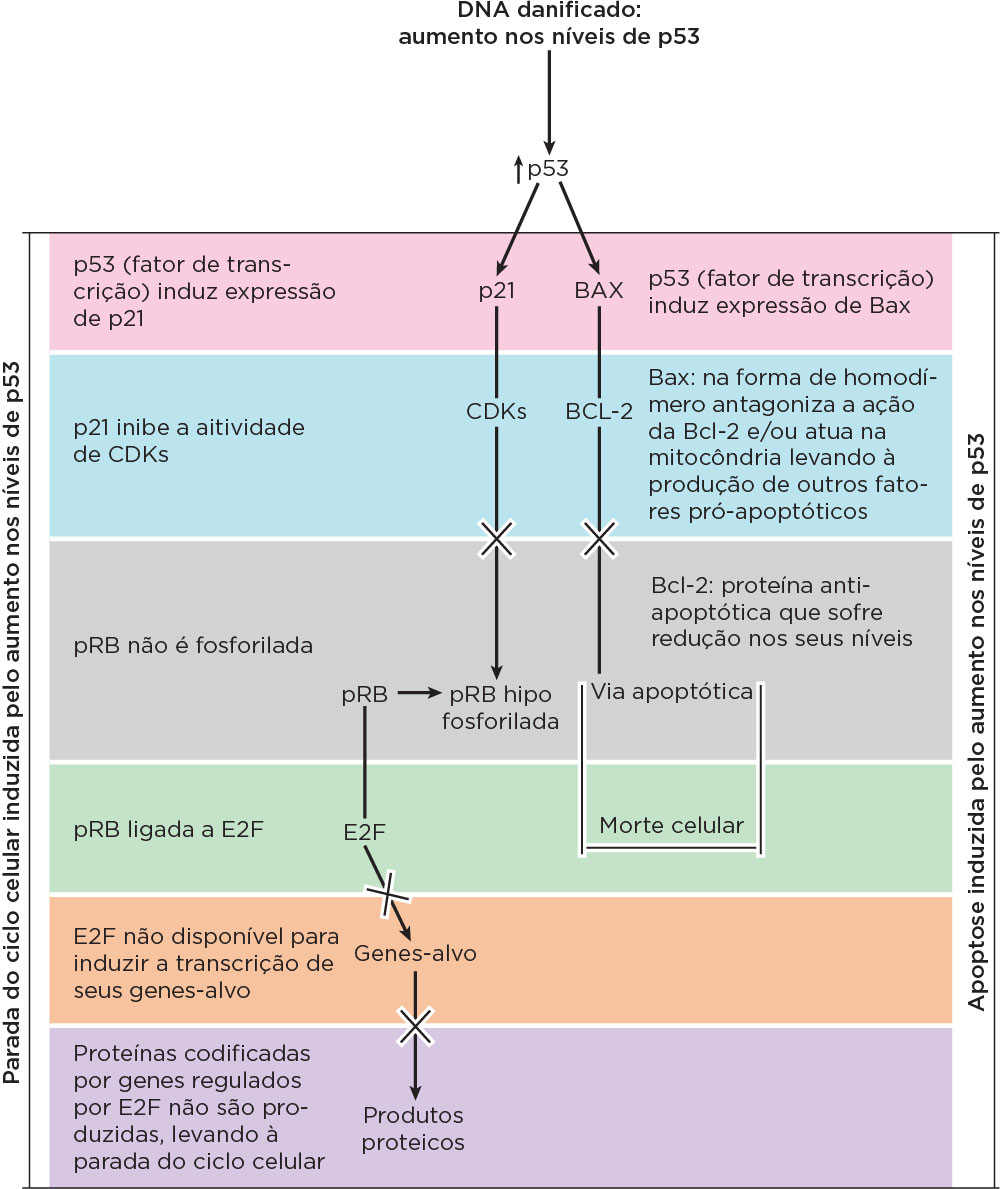
Outro importante gene supressor tumoral é o TP53 que está localizado no cromossomo 17. Quando o DNA está danificado, os níveis da proteína p53 são aumentados na célula, e o ciclo celular só passará de G1 para S se o dano for reparado. Assim, em condições normais, o nível dessa proteína é baixo, mas quando as células são tratadas com um agente que danifica o DNA, como a radiação, seus níveis são aumentados. Essa proteína atuará de duas formas como fator de transcrição: 1) pela ativação de genes envolvidos na parada no ciclo celular para que o DNA danificado seja reparado e; 2) pela ativação de genes que induzem a apoptose. No primeiro caso, a p53 liga ao promotor do gene da proteína p21. A proteína p21 é inibidora do complexo ciclina-CDK, o qual não irá fosforilar, por exemplo, a pRB. A pRB não fosforilada, não se desliga de E2F, e os genes de progressão do ciclo celular ativados por E2F não são transcritos até que o dano no DNA seja reparado. No segundo caso, a p53 ativa a transcrição de genes pro-apoptóticos como o BAX induzindo a célula à morte celular . A disfunção da p53 faz com que o ciclo celular prossiga com a mutação no DNA, permitindo sua transmissão às células descendentes e iniciando um processo neoplásico. Um exemplo de doença causada por alterações na p53 é a Síndrome de Li-Fraumeni, condição em que ocorre predisposição a desenvolver câncer em vários locais, como mama, ossos, sistema nervoso central, leucemias, entre outros. As mutações no gene TP53 são encontradas na maioria de todos os tumores. A perda de função da p53 é, portanto, uma etapa fundamental na carcinogênese.
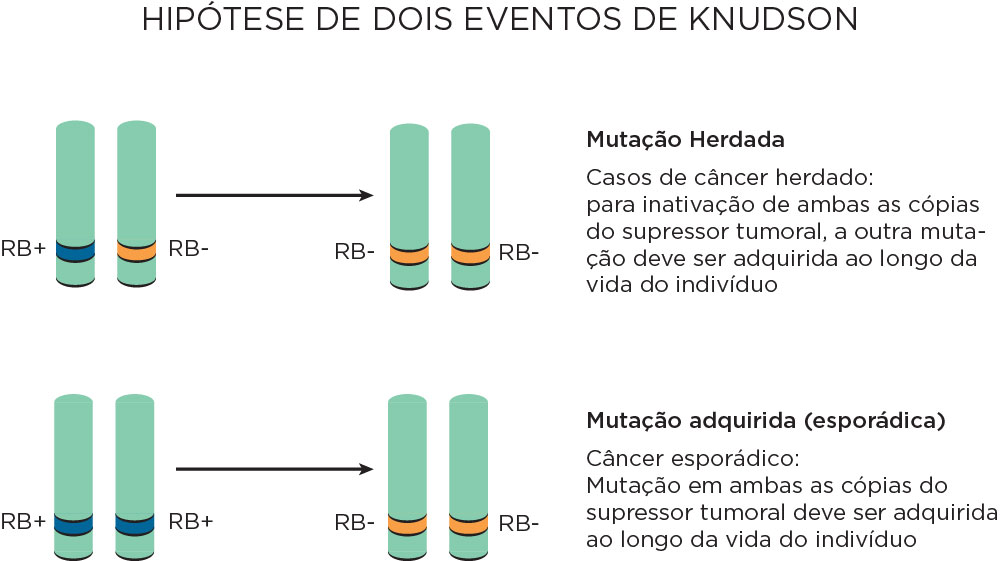
Dentre os modelos que existem para explicar a carcinogênese envolvendo genes supressores tumorais, a Hipótese dos Dois Eventos, elaborada por Alfred Knudson, em 1971 (hipótese de Knudson) é a mais aceita. Nessa hipótese, as mutações devem provocar perda de função dos dois alelos. Essas mutações têm caráter recessivo, diferente das mutações em proto-oncogenes, uma vez que um único alelo mutado não é capaz de induzir uma neoplasia. Nos tumores de caráter hereditário, quando apenas uma mutação é herdada na linhagem germinativa, a outra mutação somática deverá ser adquirida ao longo da vida. Nos tumores esporádicos, as duas mutações são somáticas e adquiridas ao longo da vida. Isso explica como algumas doenças hereditárias não se manifestam em todos os indivíduos da família, uma vez que a segunda mutação ocorre ao acaso .
Além de mutações nos genes que codificam para a pRB e p53, alterações em outros genes podem estar associadas à carcinogênese. Entre esses, estão os genes BRCA1 e BRCA2 presentes nos cromossomos 17 e 13, respectivamente. São ativados nas fases G1 e S do ciclo celular e por apresentarem uma função que parece estar associada ao reparo de danos no DNA, se os genes BRCA1 e BRCA2 estiveremse mutados, predispondo ao aparecimento de cânceres, como os de mama e de ovário. Mulheres que apresentam mutação nos genes BRCA1 e BRCA2 têm 85% de chance de desenvolver câncer de mama antes dos 70 anos de idade.