Unidade VI - Regulação da expressão gênica em eucariotos
5. Compactação do DNA Eucariótico
A associação do DNA de eucariotos às proteínas básicas histonas (H1, H2A, H2B, H3 e H4) dá ao material genético da célula uma maior condensação, formando a sua estrutura secundária. A união de duas proteínas H2A, duas H2B, duas H3 e duas H4 envolvidas por duas voltas de DNA, formam a unidade chamada de nucleossomo, que são ligados entre si pela histona H1. Assim, o enrolamento do DNA ao redor das histonas contribui para o empacotamento, reduzindo a sua extensão linear. A formação de uma estrutura mais condensada na metáfase mitótica, contendo várias voltas de DNA ao redor de várias histonas, resulta na formação de um solenoide (estrutura de 30nm) tendo em vista a associação dos nucleossomos . Em condições experimentais, o desprendimento da maioria das proteínas do DNA nos cromossomos mitóticos favorece a visualização “direta” de loops. Quando o núcleo está desprovido de proteínas, o DNA lança os loops que permanecem fixados a uma matriz proteica residual, denominada arcabouço (scaffolding) que confere aos cromossomos a sua estrutura peculiar vista na metáfase. A formação desses loops parece ser necessária para a ocorrência de transcrição.
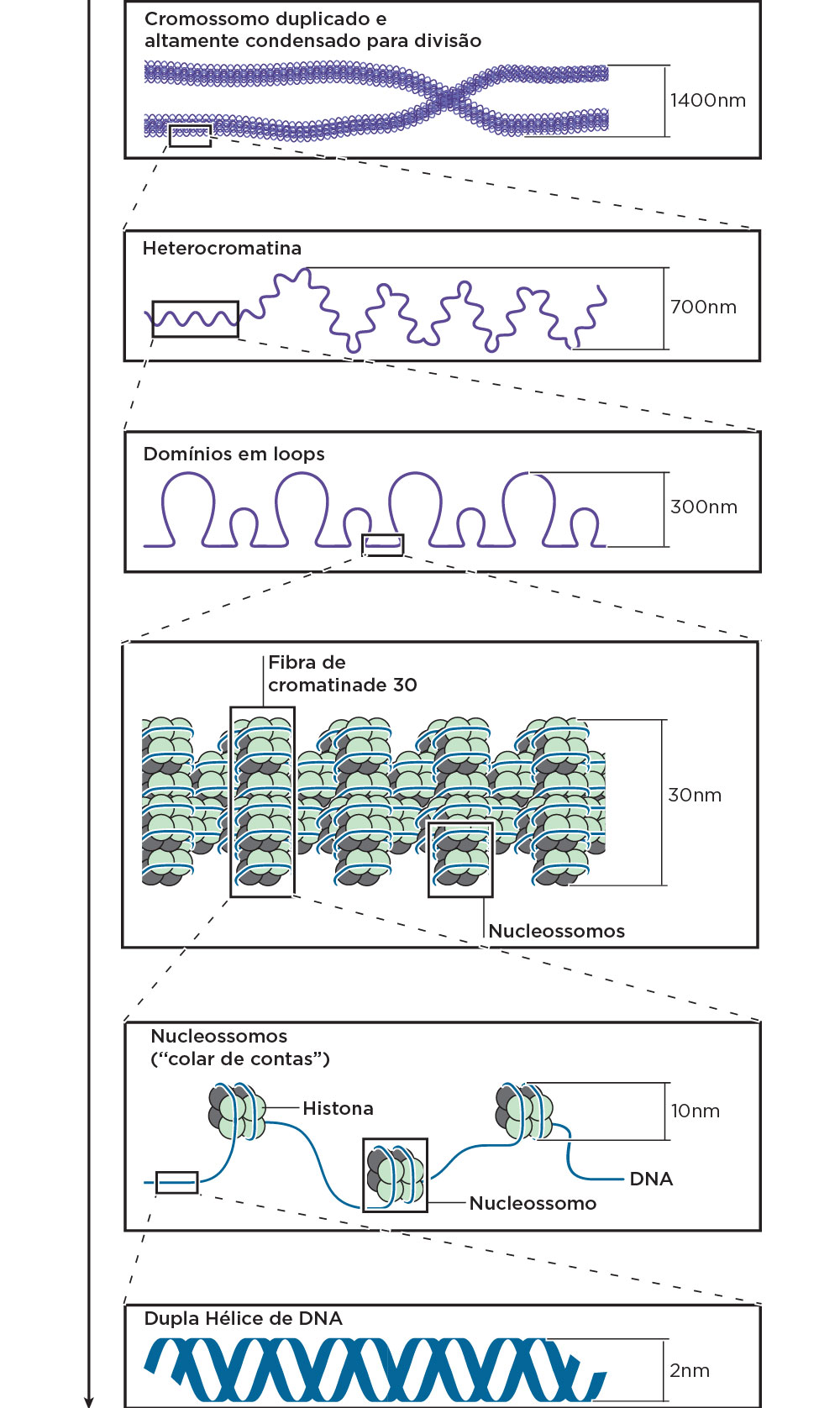
A cromatina pode ser dividida em eucromatina e heterocromatina. A eucromatina é a região no cromossomo rica em genes e transcricionalmente ativas (eu=verdadeira). Outras regiões da cromatina mais densamente condensada recebem o nome de heterocromatina, com pouca ou nenhuma atividade transcricional. Por sua vez, a heterocromatina é dividida em constitutiva, contendo regiões não expressas, como, por exemplo, algumas sequências repetidas no DNA, que desempenham papel estrutural no cromossomo, e em heterocromatina facultativa, que pode ser expressa em certos tipos celulares e em outros não.