Unidade I - Breve histórico: A descoberta do DNA como molécula da hereditariedade
Alguns Conceitos Clássicos:
- Alelo: forma alternativa de um gene que pode existir um único loco.
- Alelismo Múltiplo: conhecidos vários alelos de um gene.
- Bases Complementares: são bases que seguem um padrão de pareamento, onde A pareia com T ou U (RNA) e G com C.
- Base Nitrogenada: estrutura em anel, representada pelas pirimidinas (C,T, U) e purinas (A, G)
- Carga Genética: conjunto total de alelos deletérios contidos no genoma de um indivíduo.
- Cariótipo: conjunto cromossômico de um indivíduo visto durante a metáfase mitótica. A organização dos cromossomos por tamanho e posição do centrômero é chamada de idiograma.
- Célula Diplóide: células com dois conjuntos cromossômicos (2n).
- Células Filhas: duas células idênticas formadas a partir da divisão de uma célula.
- Célula Haplóide: célula com metade do conjunto cromossômico (n).
- Célula Somática: células não destinadas a serem gaméticas. Soma significa corpo.
- Centrômero: sequência de DNA específica que separa os braços cromossômicos eucarióticos e onde se ligam as proteínas associadas ao processo de divisão de uma célula.
- Cromossomos: o genoma contém os cromossomos: molécula longa e contínua de DNA. Cromossomos são estruturas de DNA que são vistos durante a divisão celular (Cromossomo metafásico).
- Cromossomos Autossômicos: qualquer cromossomo que não é sexual.
- Cromossomos Homólogos: cromossomos que fazem pares entre si na meiose.
- Cromossomos Sexuais: cromossomos não autossômicos cuja presença ou ausência está associada ao sexo do portador.
- DNA: ácido desoxirribonucleico, material genético básico que constitue os genes.
- Fenótipo: características manifestadas pelo indivíduo (pode mudar durante a vida): morfológicas, fisiológicas, comportamentais, outras. É determinado em sua maioria pela interação do genótipo com o ambiente.
- Gametas: célula haplóide especializada (sexual) contendo metade dos cromossomos. Exemplos: espermatozóide, óvulo e pólen.
- Genes: unidade estrutural e funcional, contituída por um segmento de DNA contendo uma sequência a ser transcrita e uma sequência regulatória que possibilita a transcrição.
- Genética: estudo dos genes e da hereditariedade. Dividida em Genética Clássia ou Mendeliana e Genética Molecular.
- Genética Molecular: estuda a estrutura e função gênica.
- Genótipo: refere-se ao conjunto de genes herdados por um indivíduo; correspondente genético da característica associada ao fenótipo; comparando ao fenótipo, o genótipo permanece “constantes” através da vida e fenótipos mudam continuamente, pois pode sofrer efeito ambiental.
- Genoma: complemento inteiro de material genético em um conjunto cromossômico. Varia de espécie para espécie. continuamente,>
- Loco (locus) ou locos (loci - plural): posição que determinado gene ocupa num par de cromossomos homólogos.
- RNA: ácido ribonucleico, material genético transcrito a partir do DNA.
- Nucleotídeo: monômero que associado forma o polímero de ácido nucleico. Constituído de um grupo fosfato, um pentose e uma base nitrogenada.
- Nucleosídeo: uma base nitrogenada ligada a uma pentose.
- Mutação: alteração na sequência de DNA, podendo produzir um gene. Cromossômica quando ocorre mudança na estrutura ou número de cromossomos na célula, cuja nomenclatura correta são alterações cromossômicas e não mutações.
- Simbologia Mendeliana para a composição genética dos indivíduos:
- Aa = heterozigoto (para um dado gene, o indivíduo porta um alelo dominante e um recessivo)
- AA = homozigoto dominante (para um dado gene, o indivíduo porta dois alelos dominantes)
- aa = homozigoto recessivo (para um dado gene, o indivíduo porta dois alelos recessivos)
- a = alelo recessivo somente se expressa quando em homozigose
Nesse item serão apresentados alguns experimentos importantes na descoberta do DNA como molécula da hereditariedade. Em 1865, o monge Gregor Mendel (1822-1884) publicou seu trabalho sobre experimentos com ervilhas que propõe as leis da hereditariedade (“leis de Mendel”) e supôs que as características hereditárias são transmitidas em unidades. O trabalho permaneceu ignorado até 1900. Em 1909, o dinamarquês Wilhelm Johannsen introduziu o termo “gene” para descrever a unidade mendeliana da hereditariedade. Ele também usou os termos “genótipo” e “fenótipo” para diferenciar as características genéticas de um indivíduo e sua aparência externa. O norte-americano Thomas Hunt Morgan, juntamente com três alunos, publicou o livro “O Mecanismo da Hereditariedade Mendeliana”, no qual relata os experimentos com drosófilas (moscas-das-frutas), e mostra que os genes estão linearmente dispostos nos cromossomos: teoria cromossômica da herança.
Paralelamente às publicações sobre heranças mendelianas, o bioquímico suíço Johann Friederich Miescher, em 1869, trabalhando com células de pus de bandagem de feridos durante a guerra da Criméia, isolou uma substância até então desconhecida, a qual ele chamou de nucleína. Essa substância continha fósforo e nitrogênio. Em 1889, seu discípulo Richard Altmann mudou o nome para ácido nucleico. Entre os anos de 1903 e 1910, o bioquímico Phoebus Aaron Levene descobriu que em alguns ácidos nucleicos também havia um açúcar do tipo ribose, e em outros um açúcar que faltava um átomo de oxigênio em um dos carbonos, a desoxirribose. Experimentos posteriores permitiram o conhecimento do monômero de nucleotídeo e cada uma das quatro bases: adenina, guanina, citosina e timina.
O inglês Frederick Griffith, em 1928, publicou resultados de experimentos de transformação utilizando penumococos encontrados em pacientes com penumonia lobar. Nesse estudo, ele utilizou duas diferentes estirpes dessas bactérias, em separado e em associação, as quais ele chamou de virulentas e não virulentas, aquecimento da cultura virulenta, centrifugação da cultura para concentração da mesma e inoculação subcutânea dessas linhagens em camundongos, a partir de, em síntese, quatro experimentos sequenciais .
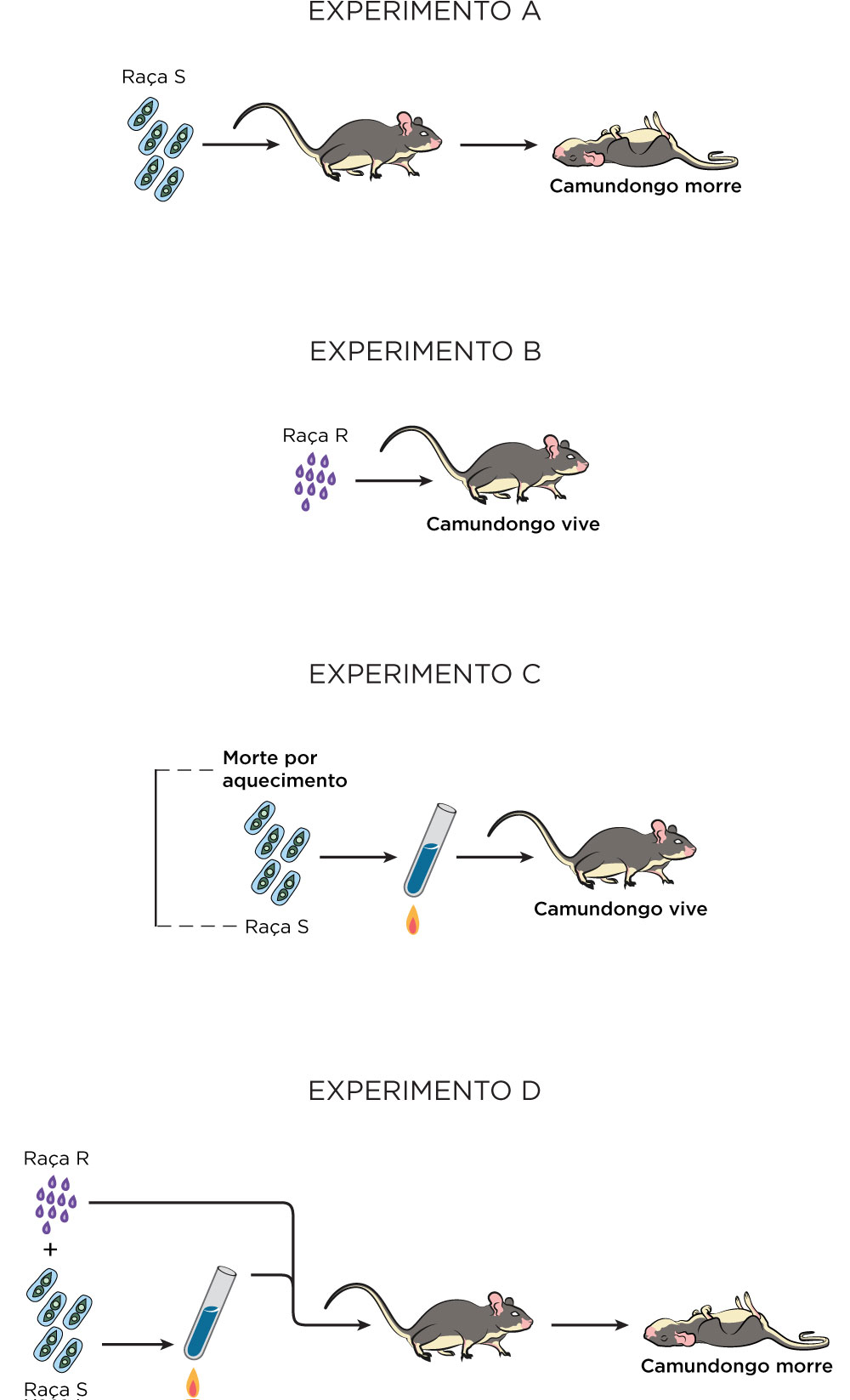
As bactérias não virulentas (pneumococos tipo R, do inglês rough, que significa rugosa) podiam matar camundongos e se tornarem encapsuladas se fossem injetadas com bactérias virulentas (tipo S, do inglês smooth, que significa lisa) mortas pelo calor. Isso mostrou que poderia haver transformações entre tipos de bactéria Streptococcus pneumoniae, que pode ser letal em camundongos. De acordo com Griffith, essa transformação seria devido a um antígeno S, que seria uma estrutura protéica específica, e quando liberado, as bactérias do tipo R poderiam utilizar para construir na sua estrutura tipo S. Ou seja, assim como todos na época, as proteínas continuavam a ser entendidas como sendo os contribuintes responsáveis pelas transmissões das características genéticas.
A natureza do princípio transformante de Griffith foi determinada pelos experimentos seguintes, publicados em 1944 por Oswald Avery, Colin MacLeod e Maclin McCarty, do Instituto Rockefeller (EUA). Dando seguimento ao trabalho na área microbiológica, com bactérias que acometem a saúde humana, esses pesquisadores trabalharam com estudos químicos de substâncias solúveis, as quais acreditavam serem importantes na patogenecidade desse microorganismos.
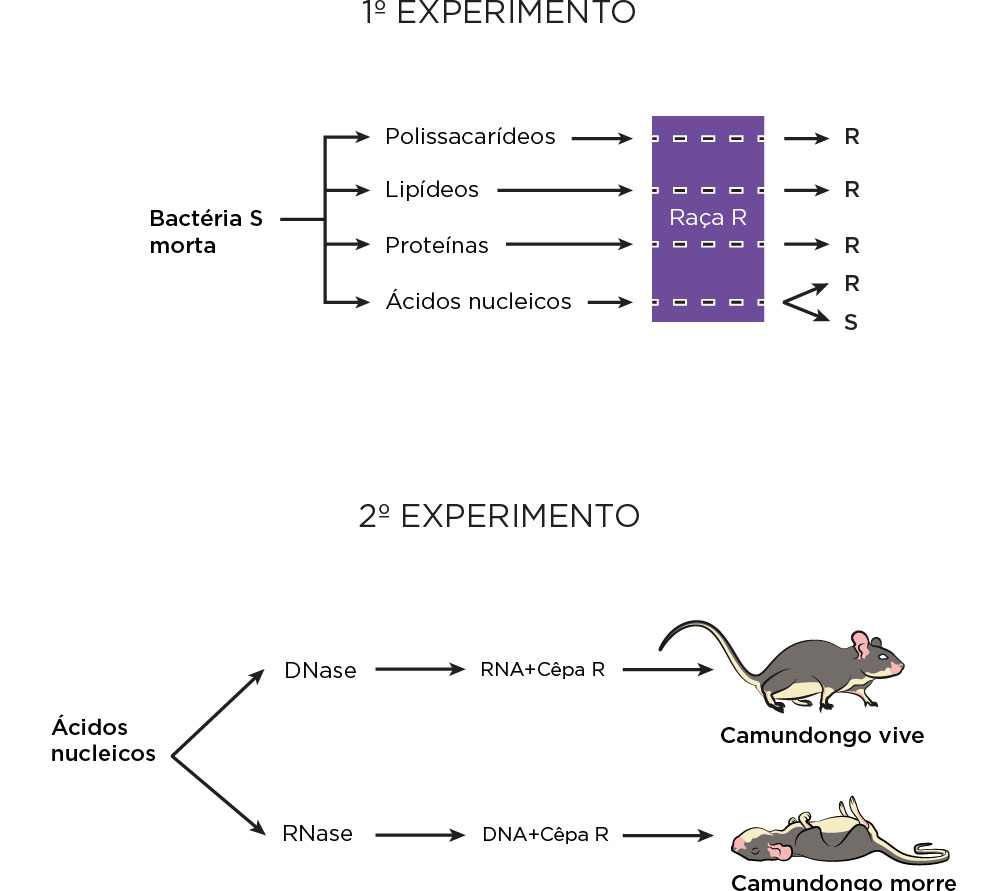
Interessados em isolar e se possível caracterizar quimicamente o suposto antígeno S, Avery, MacLeod e McCarty trabalharam com extração de frações separadas de substâncias químicas já conhecidas na época, tais como, os polissacarídeos, lipídeos, proteínas e ácidos nucleicos de bactérias do tipo S mortas pelo calor e as incubaram juntamente com as bactérias do tipo R vivas e as injetaram nos camundongos. Como resultado, somente algumas das frações contendo ácidos nucleicos foram capazes de matar os camundongos. O experimento seguinte envolveu o tratamento das frações com duas enzimas, uma que degrada o ácido ribonucleio e outra que degrada proteínas (tripsina quimiotripsina), seguida de incubação com bactérias R e inoculação nos camundongos.
Nesse estudo, esses pesquisadores observaram que a substância S não poderia ser RNA ou proteínas. Também fizeram testes combinatórios de atividade enzimática para enzimas, tais como, a fosfatase e a DNAse (chamada de despolimerase na época) em preparações de frações de tecidos como a mucosa intestinal de cachorro, rim suíno, e observaram que havia a inativação do princípio transformante sempre nas combinações que havia a enzima DNase, independente da ação das demais enzimas. Assim, nas frações de ácidos nucleicos em que restava apenas o RNA quando incubada com bactérias R e injetadas nos camundongos não levavam à morte; no entanto, a morte era causada pela incubação com o DNA.
Assim, a partir desses experimentos sequenciais, esses pesquisadores demonstraram que o ácido nucleico, do tipo desoxirribose, seria a unidade básica capaz de transformar bactérias não-patogênicas em patogênicas, e não as proteínas. No entanto, esses pesquisadores estavam cercados, na época, por químicos e físico-químicos envolvidos no estudo de proteínas, bem como por microbiologistas e virologistas com mais visibilidade no meio científico, que era difícil que esses “modestos” cientistas pudessem ter seus trabalhos bem aceitos, incluindo a percepção de que a comunidade científica da época não estava pronta para aceitar o DNA como molécula associada à hereditariedade. O próprio Avery não era geneticista e também não era conhecido pelos geneticistas, bem como a revista que escolheram para publicarem seus trabalhos não era muito conhecida ou lida por esse grupo de cientistas. Assim, continuaram acreditando que as proteínas, um dos componentes dos cromossomos, seria a molécula da hereditariedade e não o DNA, outro componente dos cromossomos; talvez, por existir traços de impurezas por proteínas que não teria sido completamente eliminado .
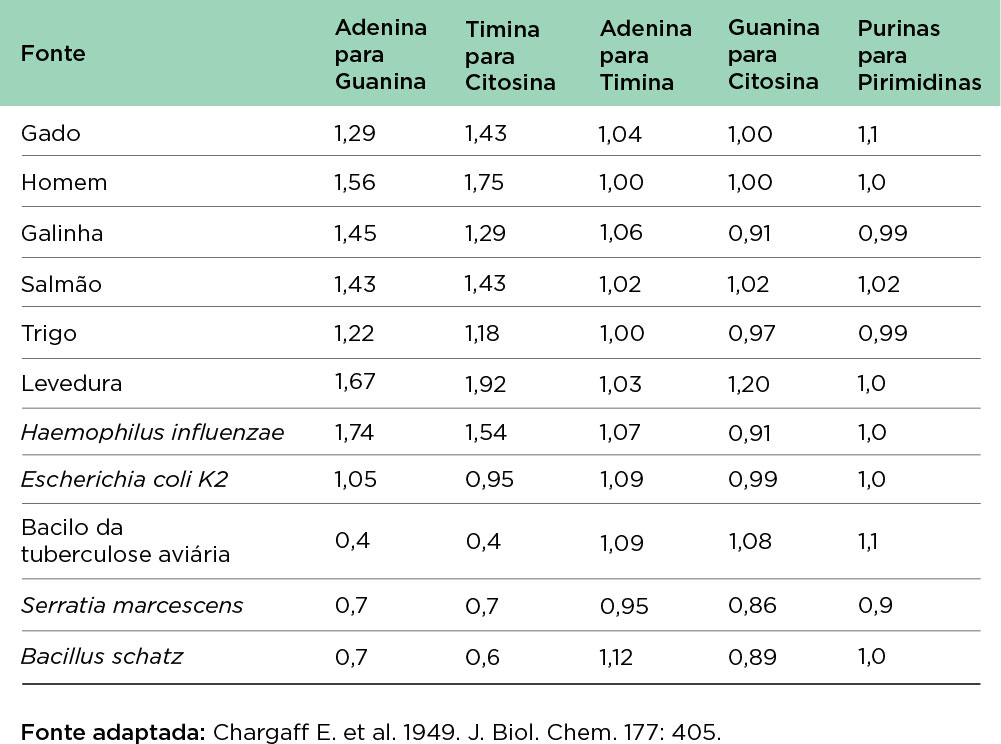
O austríaco Erwin Chargaff foi um dos poucos cientistas a reconhecer a importância dos trabalhos desenvolvidos pelo grupo de Avery. Em 1949, trabalhando com DNA de diferentes espécies e tecidos, publicou um trabalho conhecido como “regra de Chargaff” estabelecendo uma relação quantitativa entre as bases de DNA, observando uma proporção (concentrações molares) equivalente entre adenina (A) e timina (T) e o mesmo entre guanina (G) e citosina (C). Observou ainda que a relação A+T/G+C era variável entre espécies, mas a mesma entre os diferentes tecidos da mesma espécie .
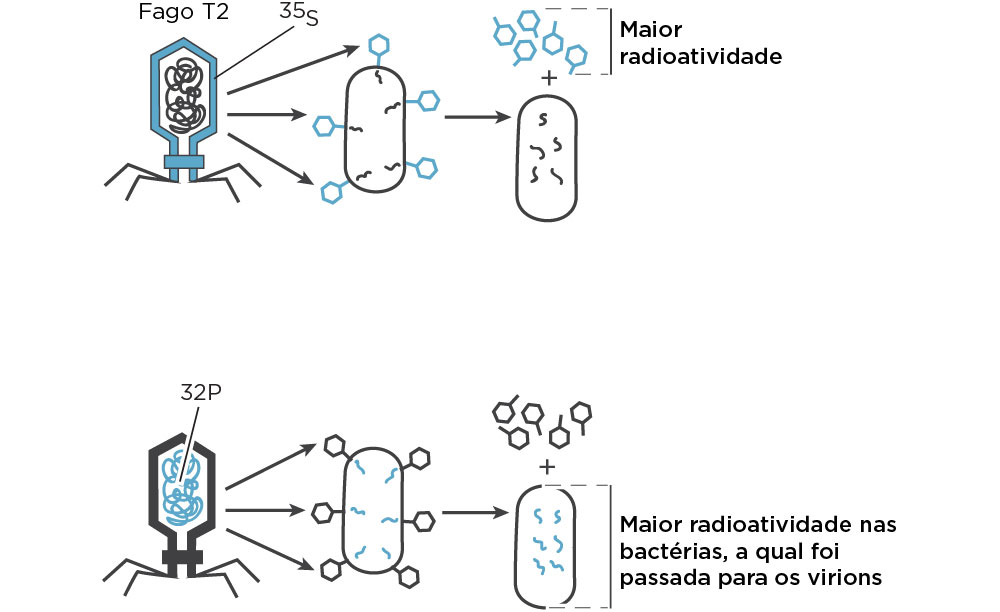
O ano de 1952 representou dois marcos importantes, tais como, a obtenção de imagens de DNA utilizando técnicas de difração de raios X pela britânica Rosalind Franklin e os experimentos dos norte-americanos Alfred Hershey e Martha Chase (Cold Spring Harbor Laboratory), os quais, utilizando marcadores radioativos, mostraram que o DNA de um bacteriófago era quem programava as células para fazer cópias do vírus e não a proteína.
O experimento de Hershey & Chase reforçou a ideia de que os genes estão contidos no DNA. Como na época já tinham o conhecimento de que o DNA continha fósforo, mas não enxofre, enquanto algumas proteínas continham enxofre, mas nenhum fósforo, esses pesquisadores marcaram o DNA pelo cultivo do fago em meio de cultura contendo o isótopo radioativo P32, enquanto a cápsula do bacteriófago foi marcada por cultivo em meio de cultura contendo S35. Por centrifugação e obtenção de duas frações, eles observaram que a maior radioatividade por P32 foi encontrada no precipitado de células bacterianas, enquanto a maior radioatividade por S35 foi encontrada no sobrenadante do tubo contendo capsídeos dos fagos, indicando que as proteínas não entravam nas bactérias. Esses dados confirmaram os achados do grupo de Avery em que o material viral injetado em E.coli foi o DNA sendo o material responsável por transferir a informação genética através das gerações, enquanto as proteínas da cápsula não eram injetadas na célula. Além disso, a prole viral continha parte de P32, mas nenhum S35. Em 1969, Hershey juntamento com Max Delbruck (coordenador do Grupo Phago) e Salvador Luria, receberam o Prêmio Nobel de Medicina .
Até esse momento, estava então, experimentalmente, demonstrado que o DNA seria a molécula da hereditariedade, no entanto, não havia ainda o entendimento de outras importantes questões, como qual a estrutura dessa molécula, como a informação genética pode ser estocada no DNA e como essa informação é passada para a prole.
Na tentativa de responder a essas questões, em 1953, os pesquisadores James Watson e Francis Crick publicaram na revista Nature dois trabalhos sobre a estrutura do DNA em dupla hélice (A Structure for Deoxyribose Nucleic Acid e Genetical Implications of the Structure of Deoxyribonucleic Acid), na qual são divulgados também outros dois artigos sobre DNA, ambos descrevendo resultados experimentais de difração de raios-X de Rosalind Franklin que eram compatíveis com a estrutura proposta por Watson e Crick. Assim, a partir de experimentos prévios realizados por outros pesquisadores, e por parcerias, revisão de trabalhos e montagem de modelos prévios não aprovados pelos pesquisadores envolvidos no estudo da estrutura do DNA, como o proposto pelo químico Linus Pauling (três hélices), Watson e Crick realizaram a montagem do quebra-cabeça.
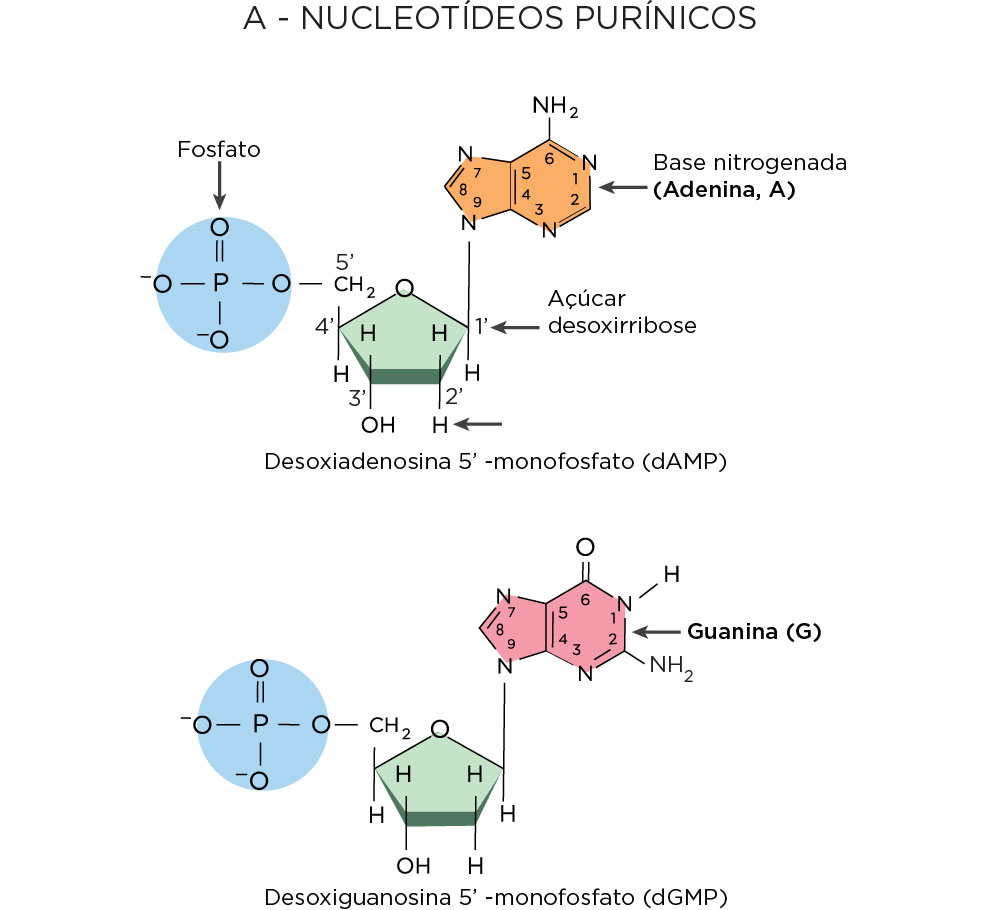
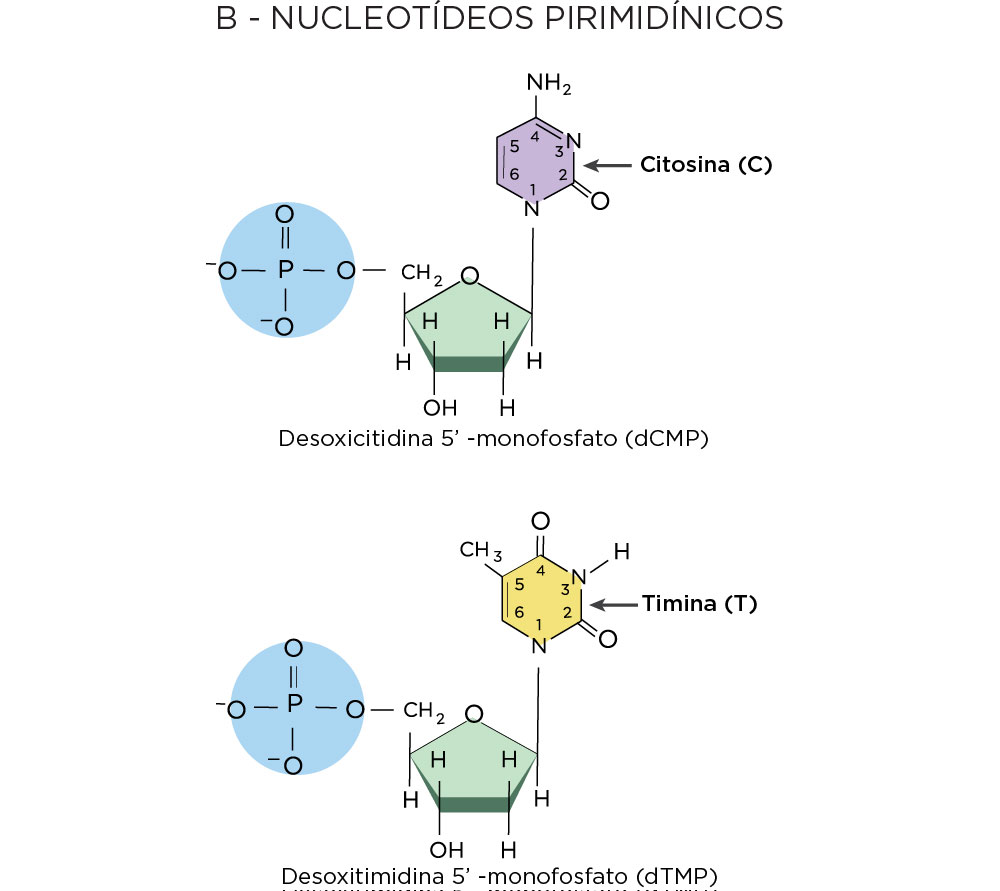
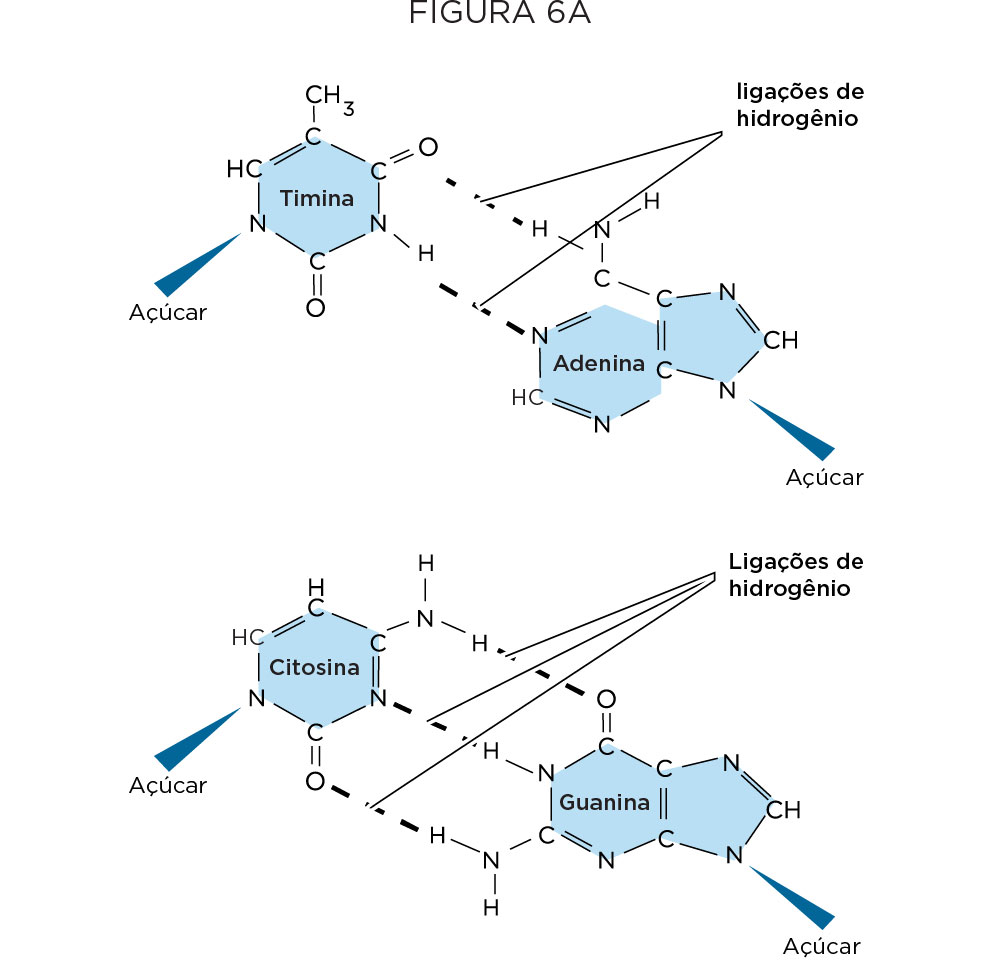
A primeira peça era o conhecimento dos nucleotídeos. De estrutura bem simples, os nucleotídeos apresentam três componentes principais, sendo um fosfato, um açúcar desoxirribose, e qualquer uma das quatro bases nitrogenadas, como bases purinas (adenina e guanina) com estrutura em dois anéis e as pirimidinas com um anel (timina e citosina) .
A segunda peça foi os resultados provenientes dos trabalhos apresentados por Chargaff sobre a composição de bases nitrogenadas do DNA de diferentes espécies.
A terceira peça veio dos dados de difração de raios-X do DNA, da física britânica Rosalind Franklin (1952). A famosa foto 51 de uma molécula de DNA mais hidratada (DNA-B) conseguida por Rosalind Franklin, bem como suas análises das medidas estruturais, foi entregue por seu chefe Raymond Gosling ao pesquisador Maurice Wilkins, do mesmo laboratório King’s College em que ela trabalhava e que também trabalhava com estrutura do DNA por difração de raios-X, o qual mostrou a Watson. A pesquisadora Franklin faleceu de câncer em 1958, possivelmente ocasionado pela exposição aos raios-X. Na elucidação da dupla hélice também foram importantes as considerações feitas pelo cristalografista Jerry Donohue a Watson & Crick de que as bases deveriam estar na sua forma ‘ceto’ e não ‘enol’ como comumente se apresentavam nos livros.
O procedimento das análises de difração de raios-X não é fácil de fazer nem de se interpretar e requer um tratamento matemático para a interpretação do padrão de pontos no filme. Os dados disponíveis sugeriram que o DNA é longo e fino e tem duas partes similares paralelas ao longo da molécula, mostrando que o DNA é uma molécula helicoidal (hélice/espiral). Assim, a partir dessa terceira peça foi possível deduzir a estrutura tridimensional do DNA, com pareamento entre purina e pirimidina compatível com os dados de raios-X; enquanto que o pareamento entre purina com purina geraria um DNA largo e no pareamento entre pirimidina com pirimidina o DNA seria muito estreito, ambos não compatíveis com os dados de raios-X.
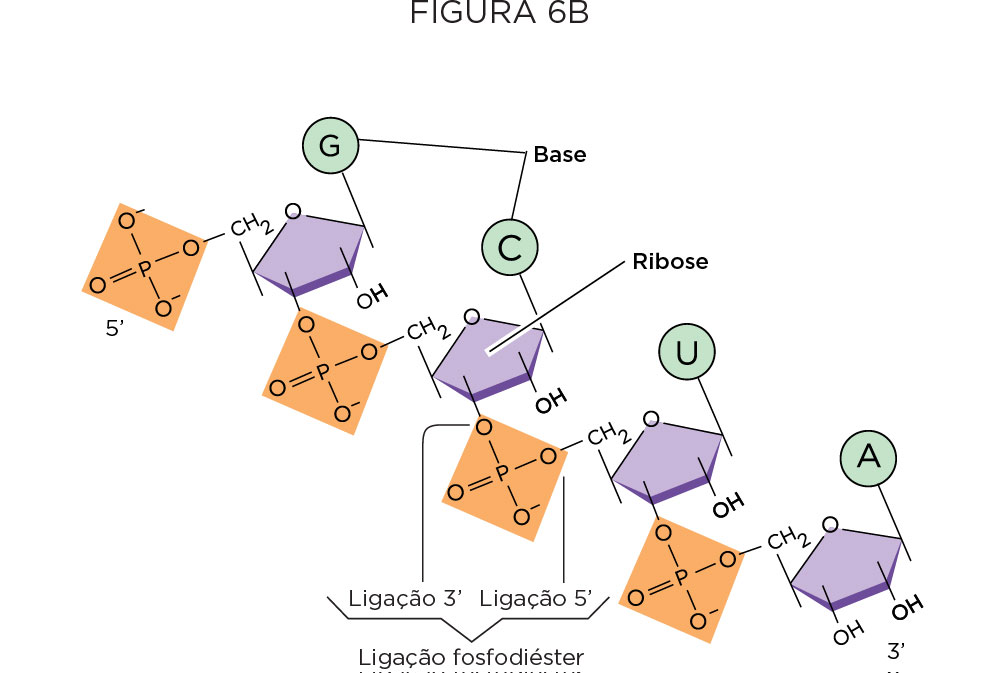
Tendo como base esses experimentos descritos anteriormente, Watson e Crick apresentaram a estrutura primária do DNA, como sendo organizado na forma de dupla hélice, sendo que cada hélice corresponde a uma cadeia de nucleotídeo disposta na polaridade inversa. Os nucleotídeos são mantidos juntos por ligações fosfodiéster entre o carbono 3’ do açúcar de um nucleotídeo e o grupo fosfato do próximo nucleotídeo, enquanto as duas hélices são mantidas juntas por pontes de hidrogênio entre as bases nitrogenadas. O pareamento de bases é específico, onde adenina (A) se pareia com timina (T), guanina (G) se pareia com citosina (C), e as ligações das purinas com as pirimidinas se dão por duas e três pontes de hidrogênio, respectivamente. A completaridade dos filamentos da dupla hélice e sua estrutura peculiar é que permite que o DNA seja o estoque da informação genética e permite que essa informação genética seja transmitida de geração a geração .
Em 1962, Watson & Crick, juntamente com Maurice Wilkins, foram agraciados com o Prêmio Nobel de Medicina, mas não a Rosalind Franklin, já que o Nobel não é conferido postumamente.
Entre outras propriedades apresentadas pela molécula de DNA, destacam-se a sua aparência altamente viscosa em soluções de DNA com pH 7,0 e na temperatura ambiente. Além disso, em altas temperaturas ou pH extremos o DNA sofre desnaturação pela ruptura das pontes de hidrogênio entre os pares de bases. A desnaturação do DNA não desfaz ligação covalente ficando, portanto, as duas fitas de DNA apenas separadas. Quando o pH e a temperatura voltam ao normal, as duas fitas de DNA espontaneamente se enrolam formando novamente o DNA dupla fita. Com base nas características de hidrofobicidade das moléculas, a estrutura de dupla hélice do DNA fica com parte hidrofílica, grupo fosfato e açúcar, localizados na parte externa da molécula e as bases nitrogenadas estão localizadas na parte interna da molécula. Ou seja, essas interações hidrofóbicas forçam as bases a se "esconderem" dentro da dupla hélice.
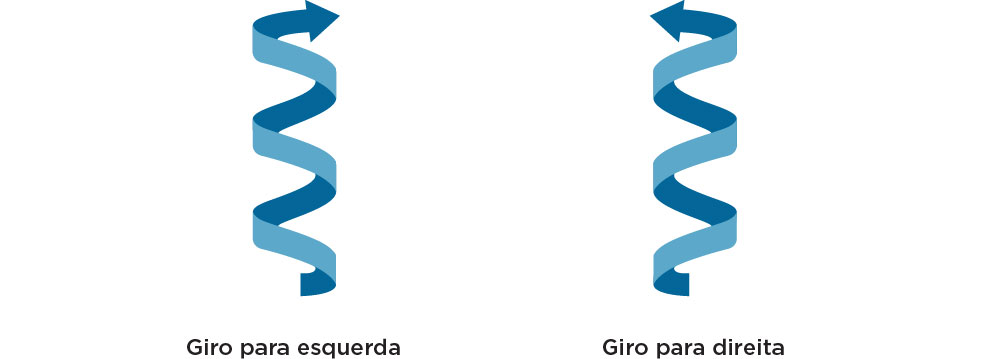
A estrutura do DNA duplex pode tomar diferentes conformações, conforme observado por Rosalind Franklin. A estrutura mais comum do DNA, semelhante ao encontrado sob condições fisiológicas e em solução, é o chamado DNA-B que se apresenta mais hidratado, com giro da hélice voltado para a direita, dupla hélice mais longa e mais fina, e 10 pares de bases são necessários para completar uma volta na hélice. O DNA assume a conformação DNA-A quando há interação DNA-RNA ou RNA-RNA, apresentando uma menor quantidade de água e, portanto, uma forma mais curta e mais grossa que o DNA-B, mas também possui giro da hélice para a direita e 11 pares de bases são necessários para completar uma volta na hélice. A relação espacial entre as duas fitas cria um sulco principal e um sulco secundário, com diferentes interações entre ligantes, tais como, proteínas. O Z-DNA tem o sentido de rotação para a esquerda, uma conformação mais alongada e mais fina do que o DNA-B, para completar uma volta na hélice são necessários 12 pares de bases e assume essa conformação em solução com altas concentrações de cátions e com sequências alternadas entre purinas e pirimidinas (por exemplo: ...GCGCGCGC...) .
De acordo com a proposição do "Dogma Central da Biologia Molecular", feita em 1958 por Francis Crick, e publicada em 1970 na revista Nature (Central Dogma of Molecular Biology), a especificidade de um fragmento de ácido nucléico depende apenas da sequência de suas bases e que essa sequência seria a chave para a disposição dos aminoácidos em uma proteína particular. Nesse sentido, propõe que o fluxo de informação vai do DNA para o RNA e depois para a proteína, e que não pode retornar.
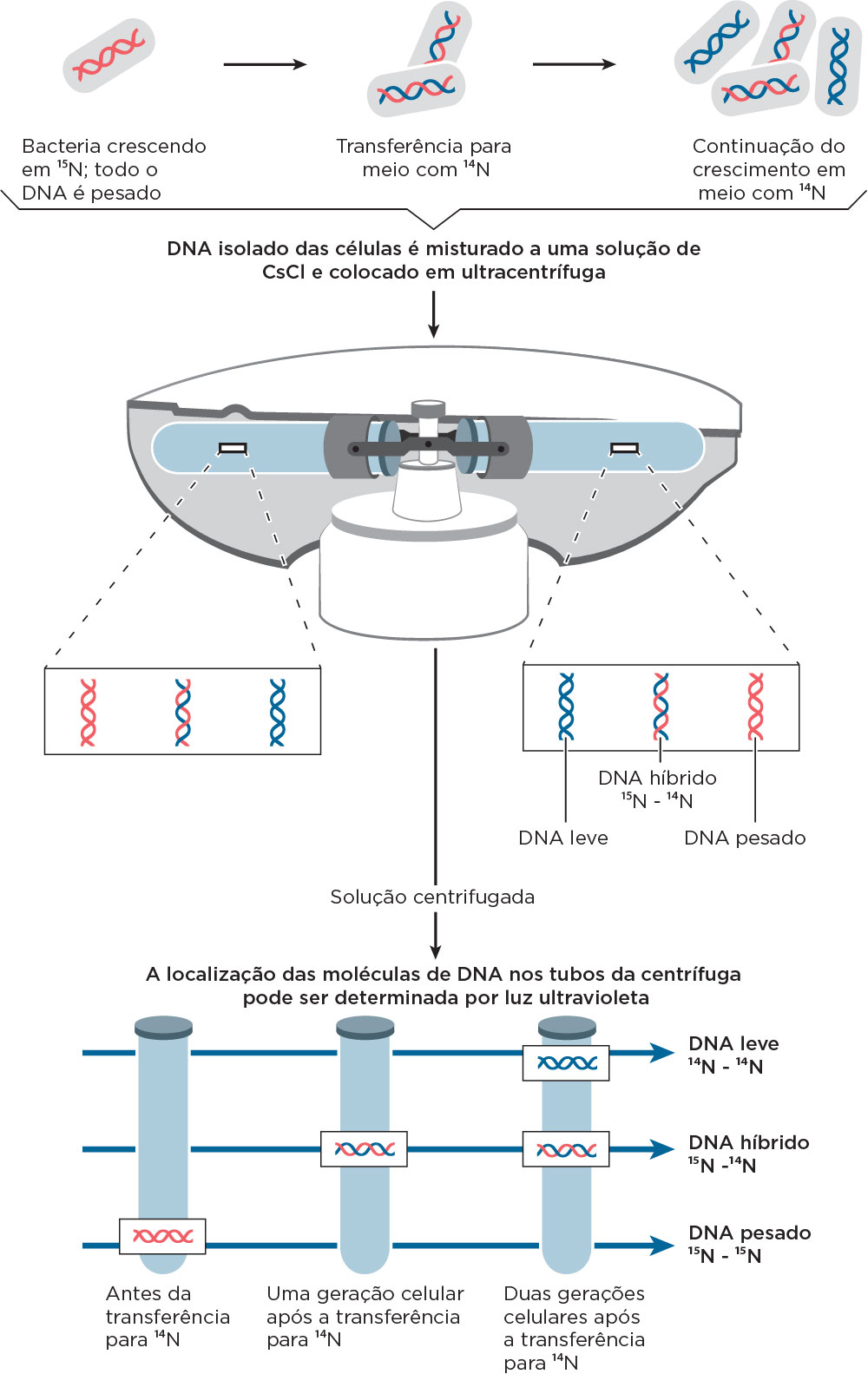
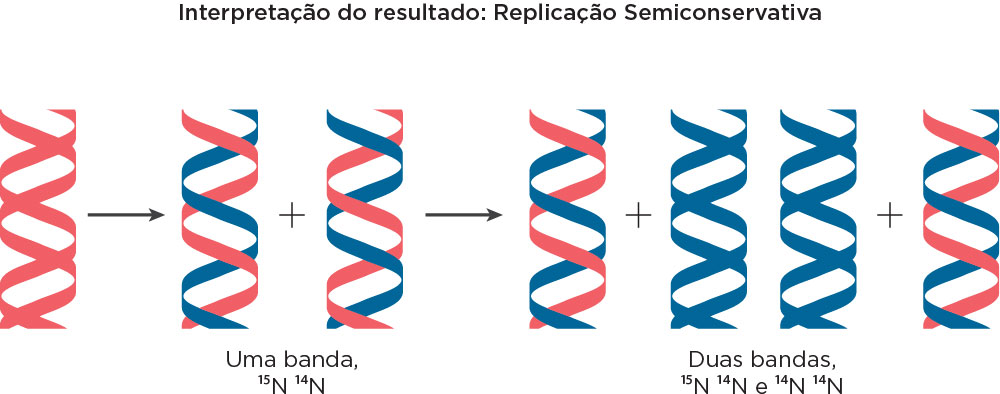
Considerando esse contexto histórico sobre o fluxo da informação genética, em 1958 os norte-americanos Mattheu Meselson e Franklin Sthal demonstraram, experimentalmente, que o DNA se replica de maneira semiconservativa, em que os dois filamentos da molécula de origem se separam e cada um deles passa a se emparelhar com um filamento novo. Na época, outros modelos foram descritos, como o modelo conservativo, em que sugeria que ambas as fitas parentais serviam como molde para a formação de duas fitas novas de DNA formando uma molécula de DNA inteiramente nova. Outro modelo, o dispersivo, sugeria que as fitas de DNA eram quebradas frequentemente em que a nova fita de DNA era formada por fragmentos novos e parentais .
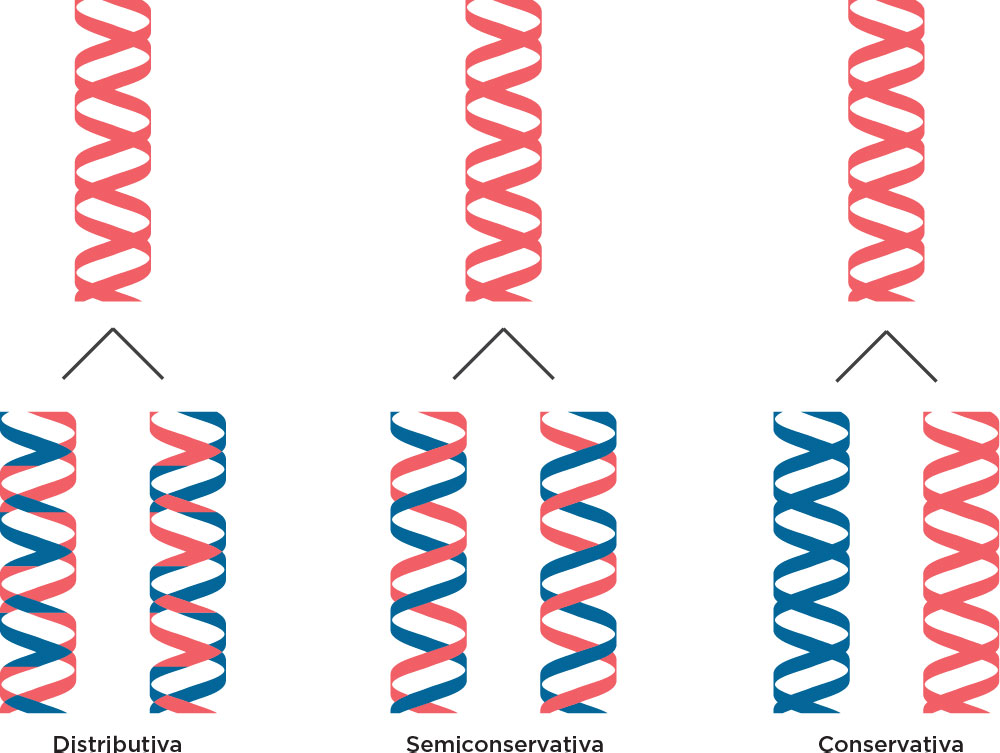
Nesse modelo, Meselson & Stahl trabalharam com células de E. coli, centrifugações, meios de cultura e isótopos radioativos. Inicialmente, as células de E. coli foram colocadas em um meio para crescimento contendo nitrogênio pesado (N15). Assim, a partir da síntese de bases nitrogenadas havia incorporação desse nitrogênio e durante o processo de replicação todo o DNA ficou marcado com este isótopo radioativo. Numa ultracentrifugação por gradiente de densidade utilizando CsCl, haveria marcação mais ao fundo do tubo. Na primeira geração de transferência dessas células para o meio contendo nitrogênio leve (N14), assim como na segunda geração também em meio contendo o N14 foram observadas marcações de densidade intermediária ao N14 e N15 (1ª. Geração) e novamente marcações de densidade intermediária entre N14 e N15 e marcações somente na densidade de N14 (2ª. Geração). Conforme pode ser observado na figura, a interpretação das bandas distintas, formadas em cada geração, indicaram um Modelo de Replicação Semiconservativa. Essas marcações poderiam ser visualizadas por luz ultravioleta .
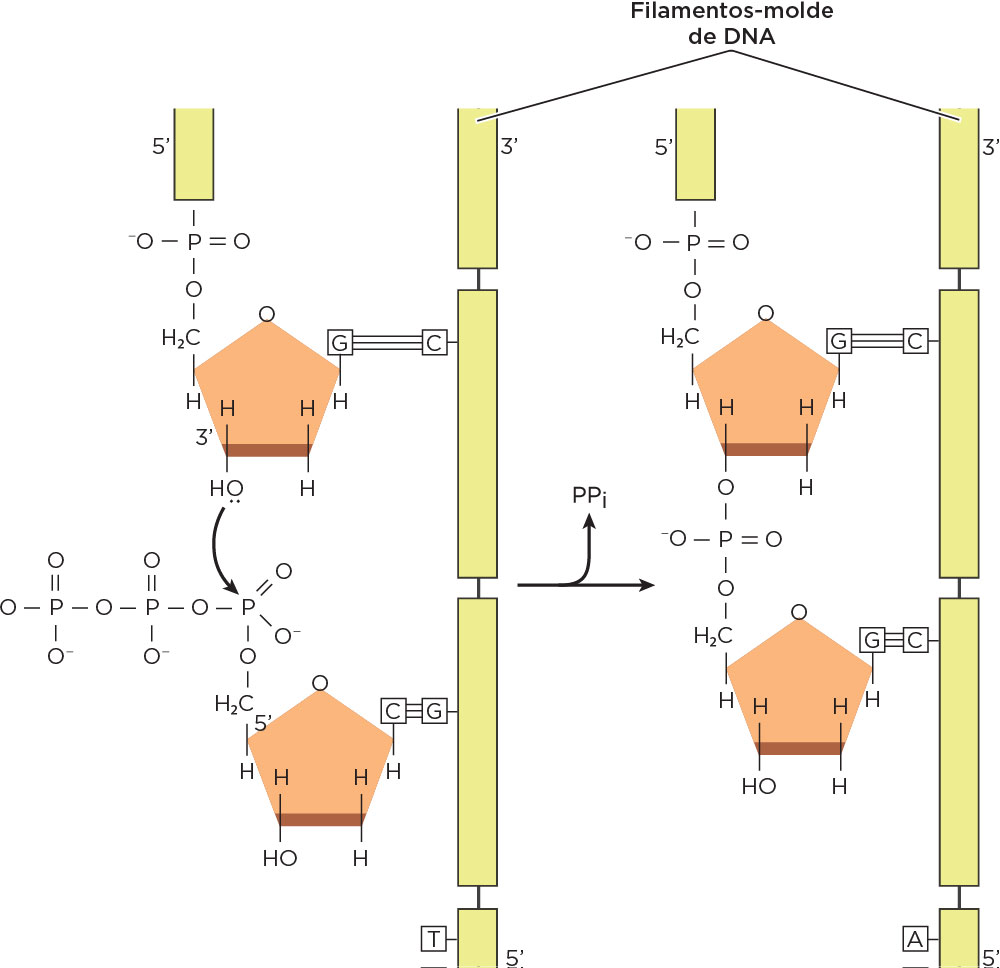
Em 1956, o norte-americano e bioquímico Arthur Kornberg identificou a polimerase, enzima que catalisa (polimeriza) a síntese de DNA utilizando extratos bacterianos livres de células, demonstrando que os nucleotídeos componentes do DNA são precursores ricos em energia (dATP, dGTP, dCTP e dTTP) e são os blocos necessários à ação dessa enzima polimerizadora . Estudos posteriores demonstraram que a enzima DNA Polimerase tipo I promove a ligação dos nucleotídeos precursores através de ligações fosfodiéster e, além disso, ela atua apenas na presença de DNA, o qual é necessário para ditar a ordem dos quatro nucleotídeos no produto polinucleotídico e na depende de uma pequena cadeia pré-existente de nucleotídeos.
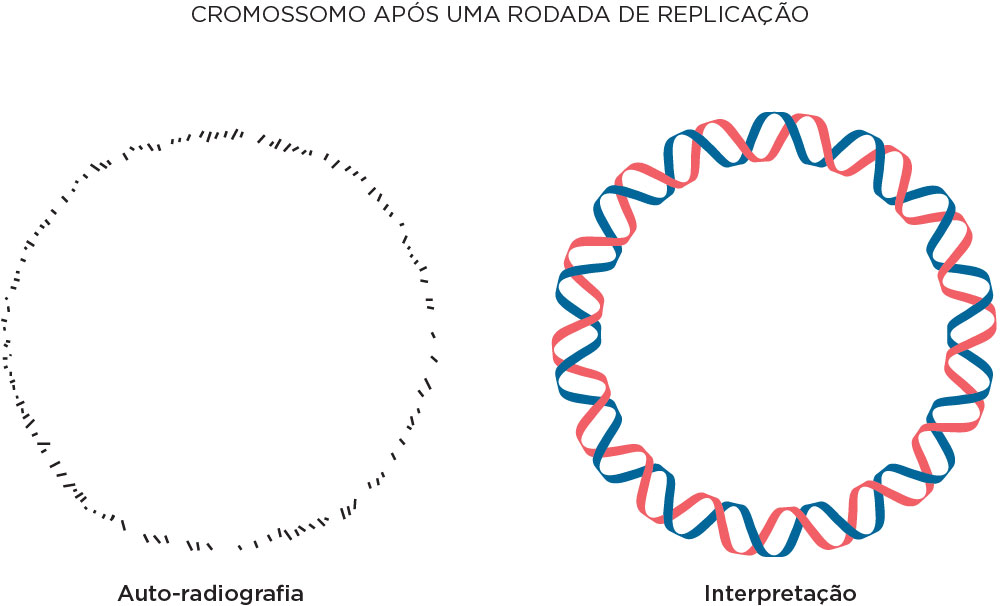
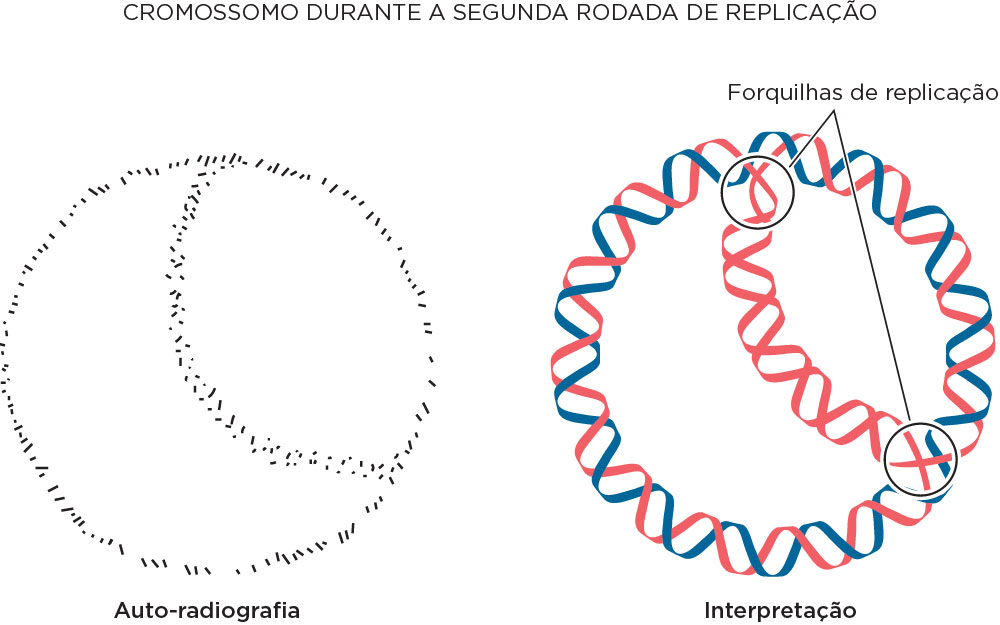
Em 1963, John Cairns demonstrou que o início da replicação do DNA se dá pela forquilha em que a dupla hélice se desenrola para produzir os dois filamentos moldes para a cópia. Utilizou timidina tritiada (3H), uma timina marcada com o isótopo radioativo de hidrogênio trítio. Com base no experimento de Meselson e Stahl, experava-se uma fita quente (radioativa) e outra fria (não radioativa). Nesse experimento, células de bactérias foram crescidas por várias gerações nesse isótopo e após lisá-la foi feita a sua transferência para um slide, análise por microscópio de luz e fez também autoradiografia. Após exposição em emulsão fotográfica de um ciclo de replicação, fez a autoradiografia obtendo uma imagem com pontos pretos em forma circular, demonstrando que o DNA bacteriano é circular. Os dados dos pontos pretos (“quentes”) foi interpretado como um filamento radioativo recém-sintetizado e outro não radioativo. Num segundo ciclo de replicação, as forquilhas foram observadas, sendo que nessa a densidade de pontos escuros permitiram a interpretação de dois filamentos radioativos na forquilha .